Aerobic training implies that the training programme is designed to improve the oxygen transport system. It is imperative during soccer match-play and training sessions that there is a good supply of oxygen to the active muscles and that these tissues have the capability to use the oxygen that is provided by the circulatory system. Aerobic training therefore has central and peripheral aspects, an effect on the cardiac output and the circulation of blood on one hand and an increased ability of the muscle to take up and utilize the oxygen that is offered.

The dimensions of the training stimulus are its duration, intensity and
frequency. The effect can be highly specific to the mode of exercise.
Improvements in aerobic fitness are reflected in the capability to sustain exercise at a given intensity for longer than was previously possible. Endurance suggests an ability to maintain exercise for a prolonged period and can be improved by focusing either on the duration or the intensity of training. Training at high intensity can entail intermittent exercise, with recovery periods intervening between the strenuous efforts. Aerobic training enhances the ability to recover quickly from strenuous activity as well as improve the capability to sustain exercise (Tomlin and Wenger, 2001).
In a soccer context the major need to raise the level of aerobic fitness applies in the pre-season period. The game itself may improve the oxygen transport system but not at a rate to achieve optimal physiological changes. For this reason the training prior to the competitive season is likely to have more formal fitness and conditioning work than at other times during the season. Gains accrued from aerobic training are likely to be less pronounced within the competitive season.

The duration of exercise that is sustainable is inversely related to the intensity at which it is performed. The longer that exercise is continued, the lower is the exercise intensity or work-rate that can be tolerated. All-out short-term exercise is fueled mainly by anaerobic sources whereas sustained endurance exercise is almost entirely supported by aerobic metabolism. An understanding of the biochemical processes involved is provided by considering the means of energy production.
Here is a chart which shows contributions of anaerobic and aerobic processes to total energy output, during maximal exercise of different duration.

AEROBIC ENERGY PRODUCTION:
Aerobic energy is produced within the muscle mitochondria by use of oxygen,
which is taken up from the blood. The substrate for this reaction may be formed through glycolysis, which refers to utilization of carbohydrates. Substrates may also be derived by catabolism of fat and, to a lesser extent, amino acids.
The net reaction of carbohydrate utilization is figured as:
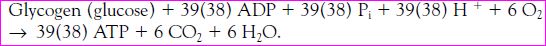
Three of the 39 ATP molecules produced are formed anaerobically. Glycogen
stored within the exercising muscles is the primary form of carbohydrate used for glycolysis, but glucose taken up in the blood from the liver also can be used. Glucose is formed in the liver from the breakdown of glycogen (glycogenolysis). It may also be formed from precursors such as glycerol, pyruvate, lactate and amino acids by means of a process known as gluconeogenesis.
The net reactions of utilizing a representative free fatty acid (such as
palmitate) are figured as:

Triglycerides (triacylglycerol) stored in the body’s adipose tissue cells and also
within the muscles form the substrates for fat oxidation. Triglycerides are broken down by lipase enzymes through a process known as lipolysis into glycerol and free fatty acids. The basic unit is one molecule of glycerol and three molecules of free fatty acids. The free fatty acids are mobilized in the bloodstream and enter the muscle fibers by diffusion where they are catabolised by the mitochondria.
Fat provides about twice as much energy per gram as does carbohydrate, and so is a good means of storing energy reserves. Oxidation of carbohydrate requires less energy than does fat and consequently the latter is the preferred fuel when exercise is at high intensity.

Fat is the preferred substrate for exercise during long training sessions and
when exercise intensity is relatively low. This form of exercise is useful for
weight-control purposes due to the so-called fat-burning effect. The relative contribution to the energy production from oxidation of carbohydrates is increased with increasing exercise intensities. Glycolysis leading to formation of lactate contributes significantly to energy production only when exercise intensities exceed about 60% of maximum oxygen uptake (VO2 max). As exercise continues, the glycogen concentration in the exercising muscles becomes progressively reduced, which leads to increases in uptake of glucose from the blood and a greater reliance on fat oxidation.
In the initial phase of exercise the production of energy by aerobic metabolism is limited due to a delay in increasing oxygen supply to the exercising muscles. Direct sources of O2 which can be used at the start of exercise comprise O2 bound to myoglobin (Mb) and hemoglobin (Hb) in blood within the muscle, and further O2 dissolved in the muscles. This local oxygen store represents only about 5% of the total energy turnover during the first 6 s of maximal exercise. The anaerobic energy production for a 30-s exercise bout contributes about 11% of the total energy turnover (Bangsbo, 1994a).
The duration as well as the intensity of the exercise bouts in an intermittent exercise programme determines the amount of lactate that accumulates both in the muscle and in the blood. Saltin and Essén (1971) kept the ratio between exercise and recovery constant at 1:2. The muscle and blood lactate concentrations were only slightly higher than at rest when the exercise time was 10 and 20 s, whereas the concentrations were considerably increased with exercise bouts of 30 and 60 s duration.
The duration of the rest periods in between the exercise bouts also affects metabolic responses during intense exercise. In a study by Margaria et al. (1969), the subjects exercised repeatedly for 10 s at an intensity that led to exhaustion after 30–40 s, when performed continuously. Blood lactate concentration increased progressively when the periods of exercise were separated by 10 s of rest, while it was only slightly elevated with 30 s of rest in between the exercise bouts.

A marked difference in muscle type recruitment has been observed between continuous and intermittent exercise protocols. While it was mainly the slow twitch(ST) that were activated during the continuous exercise, both ST and fast-twitch(FT) fibers were involved in the intermittent exercise(Essén, 1978). The different pattern of fiber type recruitment between continuous and intermittent exercise has important implications for training. By performing the training intermittently it is possible to train some muscle fibers(FT fibers) that would have been recruited only after hours of sub-maximal continuous exercise. This fact is particularly relevant for sports such as soccer in which high-intensity exercise frequently occurs. Intermittent exercise also allows a prolonged high metabolic stress without fatiguing the fibers that are recruited. Essén (1978) compared intermittent exercise with continuous exercise performed at the same power output (corresponding to VO2 max). The continuous exercise led to exhaustion within a few minutes, whereas the intermittent exercise could be sustained for 1 hour without inducing fatigue. The rate of glycogen utilization and lactate accumulation during continuous exercise at the high intensity was greater and the rate of fat oxidation was much lower than during the intermittent exercise. The intermittent protocol was a much more effective means of sparing energy reserves and extending the duration of exercise than was the continuous exercise format.


The dimensions of the training stimulus are its duration, intensity and
frequency. The effect can be highly specific to the mode of exercise.
Improvements in aerobic fitness are reflected in the capability to sustain exercise at a given intensity for longer than was previously possible. Endurance suggests an ability to maintain exercise for a prolonged period and can be improved by focusing either on the duration or the intensity of training. Training at high intensity can entail intermittent exercise, with recovery periods intervening between the strenuous efforts. Aerobic training enhances the ability to recover quickly from strenuous activity as well as improve the capability to sustain exercise (Tomlin and Wenger, 2001).
In a soccer context the major need to raise the level of aerobic fitness applies in the pre-season period. The game itself may improve the oxygen transport system but not at a rate to achieve optimal physiological changes. For this reason the training prior to the competitive season is likely to have more formal fitness and conditioning work than at other times during the season. Gains accrued from aerobic training are likely to be less pronounced within the competitive season.

The duration of exercise that is sustainable is inversely related to the intensity at which it is performed. The longer that exercise is continued, the lower is the exercise intensity or work-rate that can be tolerated. All-out short-term exercise is fueled mainly by anaerobic sources whereas sustained endurance exercise is almost entirely supported by aerobic metabolism. An understanding of the biochemical processes involved is provided by considering the means of energy production.
Here is a chart which shows contributions of anaerobic and aerobic processes to total energy output, during maximal exercise of different duration.

AEROBIC ENERGY PRODUCTION:
Aerobic energy is produced within the muscle mitochondria by use of oxygen,
which is taken up from the blood. The substrate for this reaction may be formed through glycolysis, which refers to utilization of carbohydrates. Substrates may also be derived by catabolism of fat and, to a lesser extent, amino acids.
The net reaction of carbohydrate utilization is figured as:

Three of the 39 ATP molecules produced are formed anaerobically. Glycogen
stored within the exercising muscles is the primary form of carbohydrate used for glycolysis, but glucose taken up in the blood from the liver also can be used. Glucose is formed in the liver from the breakdown of glycogen (glycogenolysis). It may also be formed from precursors such as glycerol, pyruvate, lactate and amino acids by means of a process known as gluconeogenesis.
The net reactions of utilizing a representative free fatty acid (such as
palmitate) are figured as:
Triglycerides (triacylglycerol) stored in the body’s adipose tissue cells and also
within the muscles form the substrates for fat oxidation. Triglycerides are broken down by lipase enzymes through a process known as lipolysis into glycerol and free fatty acids. The basic unit is one molecule of glycerol and three molecules of free fatty acids. The free fatty acids are mobilized in the bloodstream and enter the muscle fibers by diffusion where they are catabolised by the mitochondria.
Fat provides about twice as much energy per gram as does carbohydrate, and so is a good means of storing energy reserves. Oxidation of carbohydrate requires less energy than does fat and consequently the latter is the preferred fuel when exercise is at high intensity.
Fat is the preferred substrate for exercise during long training sessions and
when exercise intensity is relatively low. This form of exercise is useful for
weight-control purposes due to the so-called fat-burning effect. The relative contribution to the energy production from oxidation of carbohydrates is increased with increasing exercise intensities. Glycolysis leading to formation of lactate contributes significantly to energy production only when exercise intensities exceed about 60% of maximum oxygen uptake (VO2 max). As exercise continues, the glycogen concentration in the exercising muscles becomes progressively reduced, which leads to increases in uptake of glucose from the blood and a greater reliance on fat oxidation.
In the initial phase of exercise the production of energy by aerobic metabolism is limited due to a delay in increasing oxygen supply to the exercising muscles. Direct sources of O2 which can be used at the start of exercise comprise O2 bound to myoglobin (Mb) and hemoglobin (Hb) in blood within the muscle, and further O2 dissolved in the muscles. This local oxygen store represents only about 5% of the total energy turnover during the first 6 s of maximal exercise. The anaerobic energy production for a 30-s exercise bout contributes about 11% of the total energy turnover (Bangsbo, 1994a).
The duration as well as the intensity of the exercise bouts in an intermittent exercise programme determines the amount of lactate that accumulates both in the muscle and in the blood. Saltin and Essén (1971) kept the ratio between exercise and recovery constant at 1:2. The muscle and blood lactate concentrations were only slightly higher than at rest when the exercise time was 10 and 20 s, whereas the concentrations were considerably increased with exercise bouts of 30 and 60 s duration.
The duration of the rest periods in between the exercise bouts also affects metabolic responses during intense exercise. In a study by Margaria et al. (1969), the subjects exercised repeatedly for 10 s at an intensity that led to exhaustion after 30–40 s, when performed continuously. Blood lactate concentration increased progressively when the periods of exercise were separated by 10 s of rest, while it was only slightly elevated with 30 s of rest in between the exercise bouts.

A marked difference in muscle type recruitment has been observed between continuous and intermittent exercise protocols. While it was mainly the slow twitch(ST) that were activated during the continuous exercise, both ST and fast-twitch(FT) fibers were involved in the intermittent exercise(Essén, 1978). The different pattern of fiber type recruitment between continuous and intermittent exercise has important implications for training. By performing the training intermittently it is possible to train some muscle fibers(FT fibers) that would have been recruited only after hours of sub-maximal continuous exercise. This fact is particularly relevant for sports such as soccer in which high-intensity exercise frequently occurs. Intermittent exercise also allows a prolonged high metabolic stress without fatiguing the fibers that are recruited. Essén (1978) compared intermittent exercise with continuous exercise performed at the same power output (corresponding to VO2 max). The continuous exercise led to exhaustion within a few minutes, whereas the intermittent exercise could be sustained for 1 hour without inducing fatigue. The rate of glycogen utilization and lactate accumulation during continuous exercise at the high intensity was greater and the rate of fat oxidation was much lower than during the intermittent exercise. The intermittent protocol was a much more effective means of sparing energy reserves and extending the duration of exercise than was the continuous exercise format.
No comments:
Post a Comment